2 英文參考
tumor,neoplasm
腫瘤(tumor,neoplasm)是一種常見病、多發病,其中惡性腫瘤是目前危害人類健康最嚴重的一類疾病。在歐美一些國家,惡性腫瘤的死亡率僅次於心血管系統疾病而居第二位。根據我國1992年衛生事業發展情況統計公報,城市地區居民惡性腫瘤死亡率居死因第一位,爲123.92/10萬,基中男性爲151.08/10萬,女性爲95.12/10萬。農村地區居民惡性腫瘤死亡率居第死因第二位,爲101.39/10萬。其中男性爲125.79/10萬,女性爲78.85/10萬。隨着我國人口老齡化的進程,加之城市人口比例逐年增高,城鎮工業生產迅速發展,環境污染日益嚴重,吸菸等不良生活習慣相當普遍,如果不採取積極的宣傳教育措施,惡性腫瘤的危害性還將日益增加。我國最爲常見和危害性嚴重的腫瘤爲肺癌、鼻咽癌、食管癌、胃癌、大腸癌、肝癌、乳腺癌、宮頸癌、白血病及淋巴瘤等。特別是肺癌發生率近年來有明顯的增加,值得重視。這些腫瘤的病因學、發病學及其防治,均應爲我國腫瘤研究的重點。
3 腫瘤的概念
腫瘤是機體在各種致瘤因素作用下,局部組織的細胞在基因水平上失掉了對其生長的正常調控,導致異常增生而形成的新生物。這種新生物常形成局部腫塊,因而得名。
正常細胞轉變爲腫瘤細胞後就具有異常的形態、代謝和功能,並在不同程度上失去了分化成熟的能力。它生長旺盛,並具有相對的自主性,即使後來致瘤因素已不存在時,仍能持續性生長,不僅與機體不協調,而且有害無益。
機體在生理狀態下以及在炎症、損傷修復時的病理狀態下也常有組織、細胞的增生。但一般來說,這類增生有的屬於正常新陳代謝所需的細胞更新;有的是針對一定刺激或損傷的適應性反應,皆爲機體生存所需。再者,所增生的組織能分化成熟,並能恢復原來正常組織的結構和功能。而且這類增生是有一定限度的,一旦增生的原因消除後就不再繼續增生。但腫瘤性增生卻與此不同,二者有着本質上的區別。
根據腫瘤生物學特性及其對機體危害性的不同,一般分爲良性和惡性腫瘤兩大類。這種分類在腫瘤的診斷、治療和判斷預後上均有十分重要的意義。
4 腫瘤細胞的特徵
自給自足生長信號(Self-Sufficiency in Growth Signals);
抗生長信號的不敏感(Insensitivity to Antigrowth Signals);
抵抗細胞死亡(Resisting Cell Death);
潛力無限的複製能力(Limitless Replicative Potential);
持續的血管生成(Sustained Angiogenesis);
組織浸潤和轉移(Tissue Invasion and Metastasis);
避免免疫摧毀(Avoiding Immune Destruction);
促進腫瘤的炎症(Tumor Promotion Inflammation);
5 腫瘤的一般形態和結構
5.1 腫瘤的肉眼觀形態
在肉眼觀上,腫瘤的形態多種多樣,並可在一程度上反映腫瘤的良惡性。
腫瘤的數目和大小腫瘤的大小不一,通常一個,有時可爲多個。小者極小甚至在顯微鏡下才能發現,如原位癌(carcinoma in situ);大者很大,可重達數公斤乃至數十公斤。一般說,腫瘤的大小與腫瘤的性質(良、惡性)、生長時間和發生部位有一定的關係。生長於體表或大的體腔(如腹腔)內的腫瘤有時可長得很大;生長於狹小腔道(如顱腔,椎管)內的腫瘤則一般較小。大的腫瘤通常生長緩慢,生長時間較長,且多爲良性。惡性腫瘤生長迅速,短期內即可帶來不良後果,故一般不致長得很大。
腫瘤的形狀 腫瘤的形狀多種多樣,有乳頭狀、菜花狀、絨毛狀、蕈狀、息肉狀、結節狀、分葉狀、浸潤性包塊狀、瀰漫性肥厚狀、潰瘍狀和囊狀等(圖1)。腫瘤形狀上的差異一般與其發生部位、組織來源、生長方式和腫瘤的良惡性質密切相關。
腫瘤的顏色 一般腫瘤的切面多呈灰白或灰紅色,但可因其含血量的多寡、有無變性、壞死、出血,以及是否含有色素等而呈現各種不同的色調。有時可從腫瘤的色澤大致推測其爲何種腫瘤,如血管瘤多呈紅色或暗紅色,脂肪瘤呈黃色,黑素瘤多呈黑色,綠色瘤呈綠色等。

腫瘤的硬度腫瘤一般較其來源組織的硬度增大,其硬度與腫瘤的種類、瘤實質與間質的比例以及有無變性壞死等有關。如骨瘤很硬,脂肪瘤質軟;實質多於間質的腫瘤一般較軟,反之則較硬;瘤組織發生壞死時變軟,有鈣質沉着(鈣化)或骨質形成(骨化)時則變硬。
6 腫瘤的組織結構
腫瘤的組織多種多樣,但概而言之,任何一個腫瘤的組織成分都可概括爲實質和間質兩部分。
腫瘤的實質 腫瘤實質是腫瘤細胞的總稱,是腫瘤的主要成分。腫瘤的生物學特點以及每種腫瘤的特殊性都是由腫瘤的實質決定的。機體內幾乎任何組織都可發生腫瘤,因此腫瘤實質的形態也多種多樣,通常根據腫瘤的實質形態來識別各種腫瘤的組織來源(histogenesis),進行腫瘤的分類、命名和組織學診斷;並根據其分化成熟程度和異型大小來確定腫瘤的良惡性。
腫瘤的間質 腫瘤的間質成分不具特異性,起着支持和營養腫瘤實質的作用,一般系由結締組織和血管組成,有時還可有淋巴管。通常生長快的腫瘤,其間質血管多較豐富而結締組織較少;生長緩慢的腫瘤,其間質血管則較少。此外,腫瘤間質內往往有或多或少的淋巴細胞等單個核細胞浸潤,這是機體對腫瘤組織的免疫反應。近年來在腫瘤結締組織間質中除見纖維母細胞外,尚出現肌纖維母細胞(myofibroblast)。此種細胞兼具纖維母細胞和平滑肌細胞的一些形態和功能特點,細胞呈梭形。電鏡下,胞漿內除見豐富的粗麪內質網外,在細胞膜下可見平滑肌微絲。此種細胞既能產生膠原纖維,又有收縮功能。由於此種細胞增生、收縮和形成膠原纖維包繞腫瘤細胞,可能對腫瘤細胞的浸潤過程有所阻抑,並限制瘤細胞的活動和遏止瘤細胞侵入血管內或淋巴管內,從而減少播散機會。此外,此種細胞的增生還可解釋臨牀上所見乳腺癌時的乳頭回縮,食管癌及腸癌所致的管壁僵硬和狹窄。
7 腫瘤的異型性
腫瘤組織無論在細胞形態和組織結構上,都與其發源的正常組織有不同程度的差異,這種差異稱爲異型性(atypia)。腫瘤組織異型性的大小反映了腫瘤組織的成熟程度(即分化程度,在此指腫瘤的實質細胞與其來源的正常細胞和組織在形態和功能上的相似程度)。異型性小者,說明它和正常組織相似,腫瘤組織成熟程度高(分化程度高);異型性大者,表示瘤組織成熟程度低(分化程度低)。區別這種異型性的大小是診斷腫瘤,確定其良、惡性的主要組織學依據。惡性腫瘤常具有明顯的異型性。
由未分化細胞構成的惡性腫瘤也稱爲間變性腫瘤。間變(anaplasia)一詞的原意是指“退性發育”,即去分化,指已分化成熟的細胞和組織倒退分化,返回原始幼稚狀態。現已知,絕大部分的未分化的惡性腫瘤起源於組織中的幹細胞喪失了分化能力,而並非是已經分化的特異細胞去分化所致。在現代病理學中,間變指的是惡性腫瘤細胞的缺乏分化。間變性的腫瘤細胞具有明顯的多形性(pleomorphism,即瘤細胞彼此在大小和形狀上的變異)。因此,往往不能確定其組織來源。間變性腫瘤幾乎都是高度惡性的腫瘤,但大多數惡性腫瘤都顯示某種程度的分化。
7.1 腫瘤組織結構的異型性
良性腫瘤瘤細胞的異型性不明顯,一般都與其發源組織相似。因此,這種腫瘤的診斷有賴於其組織結構的異型性。例如纖維瘤的細胞和正常纖維細胞很相似,只是其排列與正常纖維組織不同,呈編織狀。惡性腫瘤的組織結構異型性明顯,瘤細胞排列更爲紊亂,失去正常的排列結構層次。例如,從纖維組織發生的惡性腫瘤—纖維肉瘤,瘤細胞很多,膠原纖維很少,排列很紊亂,與正常纖維組織的結構相差較遠;從腺上皮發生的惡性腫瘤—腺癌,其腺體的大小和形狀十分不規則,排列也較亂,腺上皮細胞排列緊密重疊或呈多層,並可有乳頭狀增生。
7.2 腫瘤細胞的異型性
良性腫瘤細胞的異型性小,一般與其發源的正常細胞相似。惡性腫瘤細胞常具有高度的特異性,表現爲以下特點:
1.瘤細胞的多形性 即瘤細胞形態及大小不一致。惡性腫瘤細胞一般比正常細胞大,各個瘤細胞的大小和形態又很不一致,有時出現瘤巨細胞(圖2)。但少數分化很差的腫瘤,其瘤細胞較正常細胞小、圓形,大小也比較一致。
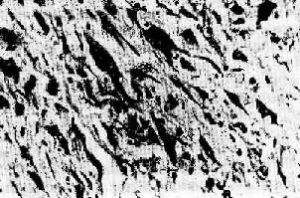
圖2 橫紋肌肉瘤
7.3 瘤細胞大小、形狀不一,並有瘤巨細胞出現
即瘤細胞核的大小、形狀及染色不一致。細胞核的體積增大(核肥大),胞核與細胞漿的比例比正常增大(正常爲1:4~6,惡性腫瘤細胞則接近1:1),核大小及形狀不一,並可出現巨核、雙核、多核或奇異形的核,核染色深(由於核內DAN增多),染色質呈粗顆粒狀,分佈不均勻,常堆積在核膜下,使核膜顯得增厚。核仁肥大,數目也常增多(可達3~5個)。核分裂像常增多,特別是出現不對稱性、多極性及頓挫性等病理性核分裂像時,對於診斷惡性腫瘤具有重要的意義。惡性腫瘤細胞的核異常改變多與染色體呈多倍體(polyploidy)或非整倍體(aneuploidy)有關。
7.4 胞漿的改變
由於胞漿內核蛋白體增多,胞漿呈嗜鹼性。並可因爲瘤細胞產生的異常分泌物或代謝產物而具有不同特點,如激素、粘液、糖原、脂質、角質和色素等。
上述瘤細胞的形態,特別是胞核的多形性常爲惡性腫瘤的重要特徵,在區別良惡性腫瘤上有重要意義。
7.5 腫瘤細胞超微結構的異型性
由於腫瘤類型不同,瘤細胞的超微結構亦有差異。一般說來,良性腫瘤細胞的超微結構基本上與其起源細胞相似。惡性腫瘤細胞根據分化的高低而表現出不同程度的異型性。總的說來,惡性腫瘤細胞的核通常絕對或相對明顯增大,核膜可有內陷或外凸,從而使核形不規則甚至形成怪形核。核仁體積增大,數目增多,形狀亦可不規則,位置往往靠邊。染色質可表現爲異染色質增加,凝集成塊狀散在或(和)邊集在核膜下。此即光鏡下所見的深染核和核膜增厚;也可表現爲常染色質爲主,即光鏡下所見染色質呈細顆粒狀的淡染核。胞漿內的細胞器常有數目減少、發育不良或形態異常,這在分化低的惡性腫瘤細胞更爲明顯。胞漿內主要見遊離的核蛋白體,而其他細胞器大爲減少。說明瘤細胞主要合成供自身生長的蛋白質,而喪失其分化的功能,這與瘤細胞生長迅速和惡性程度高有關。溶酶體在侵襲性強的瘤細胞中常有增多,它可釋放出大量水解酶,爲瘤細胞的浸潤開路。細胞間連接常有減少,這在分化低的癌更爲突出。可引起細胞間粘着鬆散,有利於細胞浸潤生長。有些在一般情況下在細胞膜表面無微絨毛的瘤細胞也可出現一些不規則微絨毛,這有利於營養物質的吸收,但有礙於細胞間的緊密接觸,從而減弱接觸抑制,有利於細胞增殖和浸潤。瘤細胞的排列結構在分化低的腫瘤也可喪失其原具有的特徵改變,例如腺癌細胞可失掉形成腺體的特徵而呈彌散排列。因此,惡性腫瘤的超微結構異型性的改變,除了瘤細胞本身的改變外,還可有排列結構的改變。
8 腫瘤細胞的代謝特點
腫瘤組織比正常組織代謝旺盛,尤以惡性腫瘤理更爲明顯。其代謝特點與正常組織相比並無質的差別,但在一定程度上反映了瘤細胞分化不成熟和生長旺盛。
8.1 核酸代謝
腫瘤組織合成DNA和RNA的聚合酶活性均較正常組織高,與此相應,核酸分解過程明顯降低,故DNA和RNA的含量在惡性腫瘤細胞均明顯增高。DNA與細胞的分裂和繁殖有關,RNA與細胞的蛋白質合成及生長有關。因此,核酸增多是腫瘤迅速生長的物質基礎。
8.2 蛋白質代謝
腫瘤組織的蛋白質合成及分解代謝都增強,但合成代謝超過分解代謝,甚至可奪取正常組織的蛋白質分解產物,合成腫瘤本身所需要的蛋白質,結果可使機體處於嚴重消耗的惡病質(cachexia)狀態。腫瘤的分解代謝表現爲蛋白質分解爲氨基酸的過程增強,而氨基酸的分解代謝則減弱,可使氨基酸重新用於蛋白質合成。這可能與腫瘤生長旺盛有關。腫瘤組織還可以合成腫瘤蛋白,作爲腫瘤特異抗原或腫瘤相關抗原,引起機體的免疫反應。有的腫瘤蛋白與胚胎組織有共同的抗原性,亦稱爲腫瘤胚胎性抗原。例如肝細胞癌能合成胎兒肝細胞所產生的甲種胎兒蛋白(AFP),此外,卵巢、睾丸含有卵黃囊結構的生殖細胞腫瘤患者血中AFP也有升高;內胚層組織發生的一些惡性腫瘤如結腸瘤、直腸癌等可產生癌胚抗原(CEA);胃癌可產生胎兒硫糖蛋白等。雖然這些抗原並無腫瘤特異性,也不是腫瘤所專有,但檢查這些抗原,並結合其他改變可幫助診斷相應的腫瘤和判斷治療後有無復發。
8.3 酶系統
腫瘤組織酶活性的改變是複雜的。除了一般在惡性腫瘤組織內氧化酶(如細胞色素氧化酶及琥珀酸脫氫酶)減少和蛋白分解酶增加外,其他酶的改變在各種腫瘤間很少是共同的,而且與正常組織比較只是含量的改變或活性的改變,並非是質的改變。例如前列腺癌的癌組織中酸性磷酸酶明顯增加,在前列腺癌伴有廣泛骨轉移時,患者血清中的酸性磷酸酶也明顯增加;骨肉瘤及肝癌時鹼性磷酸酶增加,這不但見於腫瘤組織中,還可見於病人的血清中。這些均有助臨牀診斷。
各種不同組織來源的惡性腫瘤特別是細胞分化原始幼稚者,其酶變化特點主要表現爲某些特殊功能的酶接近或完全消失,並因而導致酶譜的一致性。例如分化差的肝癌組織中有關尿素合成的特殊酶系幾乎完全消失。其酶譜乃因而趨向與其他癌組織的酶譜一致,與胚胎細胞的酶譜相似。有時還可出現通常所沒有的酶。
8.4 糖代謝
大多數正常組織在有氧時通過糖的有氧分解獲取能量,只有在缺氧時才進行無氧糖酵解。腫瘤組織則即使在氧供應充分的條件下也主要是以無氧糖酵解獲取能量。這可能是由於癌細胞線粒體的功能障礙所致,或者與瘤細胞的酶譜變化,特別是與三個糖酵解關鍵酶(己糖激酶、磷酸果糖激酶和丙酮酸激酶)活性增加和同工酶譜的改變,以及糖異生關鍵酶活性降低有關。糖酵解的許多中間產物被瘤細胞利用合成蛋白質、核酸及脂類,從而爲瘤細胞本身的生長和增生提供了必需的物質基礎。
9 腫瘤的生長與擴散
具有局部浸潤和遠處轉移能力是惡性腫瘤細胞特有的性質。並且是惡性腫瘤威脅病人健康與生命的主要原因。因此對腫瘤生長與擴散的研究已成爲腫瘤病理學的重要內容。
9.1 腫瘤生長的生物學
典型的惡性腫瘤的自然生長史可以分成幾個階段:細胞的惡性轉化→轉化細胞的克隆性增生→局部浸潤→遠處轉移。在此過程中,惡性轉化細胞的內在特點(如腫瘤細胞倍增時間)和宿主對腫瘤細胞或其產物的反應(如腫瘤血管形成)共同影響腫瘤的生長與演進。
(1)腫瘤細胞倍增時間:惡性轉化細胞的生長週期與正常細胞一樣,分爲G0、G1、S、G2和M期。多數惡性腫瘤細胞的培增時間並不是想象的那樣比正常細胞更快,而是與正常細胞相似或者長於正常細胞。
(2)生長分數(growth fraction);生長分數指腫瘤細胞羣體中處於複製階段(S+G2期)的細胞的比例。在細胞惡性轉化的初期,絕大多數的細胞處於複製期,所以生長分數很高。但是隨着腫瘤的持續生長,不斷有瘤細胞發生分化,離開複製階段的細胞越來越多,使得大多數腫瘤細胞處於G0期。即使是生長迅速的腫瘤其生長分數也只在20%左右。
(3)瘤細胞的生成與丟失:腫瘤的進行性生長及其生長速度決定於其細胞的生成大於丟失的程度。由於營養供應不足,壞死脫落以及機體抗腫瘤反應等因素的影響,在腫瘤生長過程中,有相當一部分瘤細胞失去生命力。腫瘤細胞的生成與丟失的程度共同影響着腫瘤的生長。在生長分數相對較高的腫瘤,瘤細胞的生成遠大於丟失,因此其生長速度比那些細胞生成稍超過丟失的腫瘤要快得多。
腫瘤的細胞動力學概念在腫瘤的化學治療上有重要的意義。目前幾乎所有的化學抗癌藥物均針對處於複製期的細胞。因此高生長分數的腫瘤(如高惡性的淋巴瘤)對於化療特別敏感;常見的實體瘤(如結腸癌)生長分數低,故對治療出現相對耐藥性。臨牀治療這些腫瘤的戰略是先用放射或手術治療將腫瘤縮小,使殘存的瘤細胞從G0期進入複製期後再用化療。
2.腫瘤的血管形成 臨牀與動物實驗都證明,如果沒有新生的血管供應營養,腫瘤在達到1~2mm的直徑或厚度後,即107個細胞左右將不再增大。因此誘導血管的生成能力是惡性腫瘤能生長、浸潤與轉移的前提之一。現已發現,由腫瘤細胞本身和浸潤到腫瘤組織內及其周圍的炎細胞(主要是巨噬細胞)能產生一類血管生成因子(angiogenesis factor),如纖維母細胞生長因子(fibroblastic growth factor,FGF)、血小板衍生的內皮細胞生長因子(platelet-derived endothelial cell growth factor,PD-ECGF)、轉化生長因子α(transforming growth factor-α,TGF-α)、腫瘤壞死因子α(TNF-α)、血管內皮細胞生長因子(vascular endothelial growth factor,VEGF)等。其中最具有特徵性血管生成作用的是由腫瘤細胞產生的FGF,它能通過其受體與相應的靶細胞結合。有增加內皮細胞的化學趨向性、促進血管內皮細胞分裂、毛細血管出芽生長、誘導蛋白溶解酶生成和有利於內皮細胞芽穿透基質等功能。此外,巨噬細胞產生的TNF-α也有促進內皮細胞分裂和刺激其遊走的作用。新生的毛細血管既爲腫瘤的生長提供了營養,又爲腫瘤的轉移準備了條件。因此對抑制腫瘤血管生成的研究是當前的研究熱點之一。
3.腫瘤的演進與異質化 惡性腫瘤在生長過程中變得越來越富有侵襲性的現象稱爲腫瘤的演進(progression),包括生長加快、浸潤周圍組織和遠處轉移等。這些生物學現象的出現與腫瘤細胞的不同亞克隆在侵襲能力、生長速度、對激素的反應、對抗癌藥的敏感性等方面的差異—腫瘤的異質性(heterogeneity)有關。產生這種現象的原因是在腫瘤的生長過程中,可能有附加的基因突變(詳見第十五節)作用於不同的瘤細胞,使得瘤細胞的亞克隆獲得不同的特性。例如需要較多生長因子的亞克隆可因生長因子缺乏而不能生長,而有些需要較少生長因子的亞克隆在此時即可生長;機體的抗腫瘤反應可殺死那些具有較高的抗原性的亞克隆,而抗原性低的亞克隆則可以躲過機體的免疫監視。由於這些選擇,腫瘤在生長過程中能保留那些適應存活、生長、浸潤與轉移的亞克隆。這就是腫瘤的異質化。
9.2 腫瘤的生長方式和擴散
各種腫瘤的生長速度有極大的差異,主要決定於腫瘤細胞的分化成熟程度。一般來講,成熟程度高、分化好的良性腫瘤生長較緩慢,幾年甚至幾十年。如果其生長速度突然加快,就要考慮發生惡性轉變的可能。成熟程度低、分化差的惡性腫瘤生長較快,短期內即可形成明顯的腫塊,並且由於血管形成及營養供應相對不足,易發生壞死,出血等繼發改變。
(1)膨脹性生長:這是大多數良性腫瘤所表現的生長方式。由於這種瘤細胞生長緩慢,不侵襲周圍正常組織,隨着腫瘤體積的逐漸增大,有如逐漸膨脹的氣球,向四周組織推擠。因此腫瘤往往呈結節狀,周圍常有完整的包膜,與周圍組織分界清楚(圖7-3)。位於皮下者臨牀觸診時可以推動,容易手術摘除,摘除後也不易復發。雖這種生長方式的腫瘤對局部器官、組織的影響主要爲擠壓或阻塞,一般均不明顯破壞器官的結構和功能。
(2)外生性生長:發生在體表、體腔表面或管道器官(如消化道,泌尿生殖道等)表面的腫瘤,常向表面生長,形成突起的乳頭狀、息肉狀、蕈狀或菜花狀的腫物。這種生長方式稱爲外生性生長。良性腫瘤和惡性腫瘤都可呈外生性生長。但惡性腫瘤在外生性生長的同時,其基底部往往也呈浸潤性生長,又由於其生長迅速,血液供應不足,這種外生性腫物容易發生壞死脫落而形成底部高低不平、邊緣隆起的癌性潰瘍。
(3)浸潤性生長:爲大多數惡性腫瘤的生長方式。瘤細胞分裂增生,侵入周圍組織間隙、淋巴管或血管內,如樹根之長入泥土,浸潤並破壞周圍組織。因而此類腫瘤沒有包膜,與鄰近的正常組織緊密連接在一起而無明顯界限(圖4)。臨牀觸診時,腫瘤固定不活動。手術切除這種腫瘤時,切除範圍比肉眼所見腫瘤範圍爲大,因爲這些部位也可能有腫瘤細胞的浸潤。

癌細胞形成大小、形狀不一的細胞巢,浸潤於管壁組織(包括肌層)間隙內
具有浸潤性生長的惡性腫瘤,不僅可以在原發部位繼續生長、蔓延(直接蔓延)而且還可以通過多種途徑擴散至身體其他部位(轉移)。
(1)直接蔓延:隨着腫瘤的不斷長大,瘤細胞常常連續不斷地沿着組織間隙、淋巴管、血管或神經束衣侵入並破壞鄰近正常器官或組織,並繼續生長,稱爲直接蔓延。例如晚期子宮頸癌可蔓延至直腸和膀胱;晚期乳腺癌可穿過胸肌和胸腔甚至達肺。
(2)轉移(metastasis):瘤細胞從原發部位侵入淋巴管、血管或體腔,被帶到他處而繼續生長,形成與原發瘤同樣類型的腫瘤,這個過程稱爲轉移。所形成的腫瘤稱爲轉移瘤或繼發瘤。良性腫瘤不轉移,只有惡性腫瘤纔可能發生轉移。常見的轉移途徑有以下幾種:
1)淋巴道轉移:瘤細胞侵入淋巴管(圖5)後,隨淋巴流首先到達局部淋巴結。例如乳腺外上象限發生的乳腺癌首先到達同側腋窩淋巴結;肺癌首先到達肺門淋巴結。瘤細胞到達局部淋巴結後,先聚集於邊緣竇,以後生長繁殖而累及整個淋巴結(圖6),使淋巴結腫大,質地變硬,切面常呈灰白色。有時有轉移的淋巴結由於瘤組織侵出被膜而互相融合成團塊。局部淋巴結發生轉移後,可繼續轉移至下一站的其他淋巴結,最後可經胸導管進入血流再繼發血道轉移。


2)血道轉移:瘤細胞侵入血管後可隨血流到達遠隔器官繼續生長,形成轉移瘤。由於動脈壁較厚,同時管內壓力較高,故瘤細胞多經小靜脈入血。少數亦可經過淋巴管入血。血道轉移的運行途徑與血栓栓塞過程相同,即侵入體循環靜脈的腫瘤細胞經右心到肺,在肺內形成轉移瘤,例如骨肉瘤等的肺轉移;侵入門靜脈系統的腫瘤細胞,首先形成肝內轉移,例如胃、腸癌的肝轉移等;侵入肺靜脈的腫瘤細胞或肺內轉移瘤通過肺毛細血管而進入肺靜脈的瘤細胞,可經左心隨主動脈血流到達全身各器官,常見轉移腦、骨、腎及腎上腺等處。因此,這些器官的轉移瘤常發生在肺已有轉移之後。此外,侵入胸、腰、骨盆靜脈的腫瘤細胞,也可以通過吻合支進入脊椎靜脈叢(Batson脊椎靜脈系統),例如前列腺癌就可通過此途徑轉移到脊椎,進而轉移到腦,這時可不伴有肺的轉移。
血道轉移雖然可見於許多器官,但最常見的是肺,其次是肝。故臨牀上判斷有無血道轉移,以確定患者的臨牀分期和治療方案時,作肺部的X線檢查及肝的超聲等影像學探查是非常必要的。轉移瘤在形態上的特點是邊界清楚並常爲多個散在分佈的結節,且多接近器官的表面(圖7-7)。位於器官表面的轉移瘤,由於瘤結節中央出血、壞死而下陷,可形成“癌臍”。
3)種植性轉移:體腔內器官的腫瘤蔓延而於器官表面時,瘤細胞可以脫落並象播種一樣,種植在體腔和體腔內各器官的表面,形成多數的轉移瘤。這種轉移的方式稱爲種植性轉移或播種。種植性轉移常見於腹腔器官的癌瘤。如胃癌破壞胃壁侵及漿膜後,可種植到大網膜、腹膜、腹腔內器官表面甚至卵巢等處。肺癌也常在胸腔內形成廣泛的種植性轉移。漿膜腔的種植性轉移每伴有漿液性血性積液。這是由於漿膜下淋巴管或毛細血管被癌栓阻塞,或漿膜受癌的刺激,使其內毛細血管的通透性增加而致滲出增多,並由於血管被癌細胞破壞而引起出血這故。抽吸積液作細胞學檢查常可查見癌細胞。腦部的惡性腫瘤,如小腦的髓母細胞瘤(medulloblastoma)亦可經腦脊液轉移到腦的其他部位或脊髓,形成種植性轉移。值得注意的是,手術也可能造成種植轉移,應注意儘量避免。

圖7 肺內的血道轉移性癌
(1)局部浸潤:惡性腫瘤局部浸潤的機制目前尚未十分明瞭,但已知是一個由一系列步驟組成的複雜過程。前面提到的浸潤能力強的瘤細胞亞克隆的出現和腫瘤血管形成對此都起着重要的作用。局部浸潤發生時,由細胞粘附分子(cell adhesion molecules)介導的腫瘤細胞彼此之間的粘附力減弱和瘤細胞與基質的附着力增加是近年來關於侵潤機制研究的重點。已有資料顯示,在動物高浸潤性的腫瘤細胞株,其瘤細胞表面的一種粘附分子——上皮粘連素(E—cadherin)的表達減少,而如將編碼上皮粘連素DNA插入到瘤細胞基因組中,則可使其喪失轉移和浸潤能力。瘤細胞彼此分散才能侵入細胞外基質(extracellular matrix,ECM)。細胞外基質在機體內分隔上皮組織和結締組織,包括基底膜和間質性結締組織,主要是由膠原、糖蛋白和蛋白多糖組成。正常上皮細胞與基質的結合是通過存在於上皮細胞膜表面的整合素(integrin)的粘附分子與存在於基質中的其配體的結合來實現的。
癌細胞的ECM主要成成分——基底膜的侵襲是一主動過程,可分爲三個步驟:①癌細胞附着於基底膜(attachment):正常上皮細胞具有的一種整合素——層粘連蛋白(laminin,LN)的受體,只分佈在細胞的基底面,能與基底膜的LN分子結合而使上皮細胞附着。而癌細胞則有更多的LN受體,分佈於癌細胞的整個表面,使癌細胞更容易與基底膜粘附。例如已發現,人的侵潤性乳腺癌細胞與LN的結合能力爲正常或良性乳腺上皮細胞的50倍,這類病人發生淋巴結轉移的機會大大高於LN受體較少的乳腺癌病人。纖維粘連蛋白(fibronectin,FN)也是基底膜的成分,可與上皮細胞表面的一種整合素——FN受體結合而使細胞附着於基底膜。癌細胞FN受體表達增加和其侵襲性之間也存在於與LN類似的關係;②細胞外基質的降解(degradation);在癌細胞與基底膜緊密接觸4~8小時後,ECM的成分,如LN、FN、蛋白多糖和膠原纖維(Ⅳ型)可被癌細胞直接分泌的蛋白溶解酶(包括Ⅳ型膠原酶、尿激酶型胞漿素原活化物、組織蛋白酶D等)溶解,使基底膜產生局部的缺損。癌細胞也可誘導宿主細胞(如纖維母細胞)產生蛋白酶,使ECM溶解。Ⅳ型膠原酶是一種金屬蛋白酶,能分解上皮和血管基底膜的Ⅳ型膠原纖維,已有報告指出在乳腺癌和胃癌細胞有這種酶的過度表達。動物實驗還發現Ⅳ型膠原酶的抑制劑可以大大減少轉移的發生;③癌細胞的移出(migration);癌細胞通過被溶解的基底膜缺損處游出是藉助於自身的阿米巴運動。近來發現腫瘤細胞的衍生的細胞激肽,如自分泌移動因子(autocrine motility factor)可介導瘤細胞的移動。基質成分(如膠原、LN)的降解產物和某些生長因子(如胰島素樣生長因子Ⅰ和Ⅱ)對癌細胞有化學趨向性。癌細胞穿過基底膜後,重複上述步驟進一步溶解間質性的結締組織,在間質中移動。到達血管壁時,可以以同樣方式穿過血管的基底膜進入血管(圖7-8)。
(2)血行播散:進入血管的癌細胞形成新的轉移竈的可能性小於千分之一,因單個癌細胞進入血管後絕大多數爲機體的免疫細胞消滅。但被血小板凝集成團的癌細胞形成的瘤栓則不易被消滅,並可與形成栓塞處的血管內皮細胞粘附,然後以前述機制穿過血管內皮和基底膜,形成新的轉移竈(圖7-8)。由於腫瘤的異質化而選擇出的高侵襲性的瘤細胞亞克隆,尤其容易形成廣泛的血行播散。
轉移的發生不是隨機的。早在1889年Paget在對700多側乳腺癌的轉移進行分析後,就發現有明顯的器官傾向性,並提出有名的“種子和土壤”學說。血行轉移的位置和器官分佈,在某些腫瘤具有特殊的親和性,如肺癌易轉移到腎上腺和腦;甲狀腺癌、腎癌和前列腺癌易轉移到骨;乳腺癌常轉移到肺、肝、骨、卵巢和腎上腺等。產生這種現象的原因還不清楚,可能是由於這些器官的血管內皮上有能與進入血循環的癌細胞表面的粘附分子特異性結合的配體(如血管細胞粘附分子),或者由於靶器官能夠釋放某些吸引癌細胞的化學吸引物質(如胰島素樣生長因子Ⅰ和Ⅱ)。此外,轉移瘤在某些組織或器官中不易形成,也可能與這些器官或組織的環境不適合腫瘤的生長有關。如脾雖然血液循環豐富但轉移癌少見,可能是因爲脾是免疫器官;橫紋肌轉移瘤很少,可能是因爲肌肉經常收縮使瘤細胞不易停留或肌肉內乳酸含量過高,不利於腫瘤生長。
(3)腫瘤轉移的分子遺傳學:目前尚發現一個單獨的轉移基因,但已發現一種腫瘤抑制基因——nm23的表達水平與腫瘤的侵襲和轉移能力之間存在有意義的關係。在小鼠模型中,nm23的表達高者具有低轉移性;nm23表達低10倍者伴有高轉移。人類的nm23基因定位於第17位號染色體。在侵襲性強的腫瘤中nm23基因丟失。臨牀上對人乳腺癌的觀察發現,淋巴結轉移少於三個者,nm23蛋白表達水平高;而有廣泛轉移者nm23蛋白表達的水平一般均低。如能將nm23蛋白作爲標記物來預測轉移並且在治療上用於抑制轉移,將是腫瘤治療的一個突破。現此問題正在積極研究中。
9.3 腫瘤的分級與分期
腫瘤的分級(grading)和分期(staging)一般都用於惡性腫瘤。惡性腫瘤是根據其分化程度的高低、異型性的大小及核分裂像的多少來確定惡性程度的級別。近年來較多的人傾向於用簡明的、較易掌握的三級分級法,即Ⅰ級爲分化良好的,屬低度惡性;Ⅱ級爲分化中等的,屬中度惡性;Ⅲ級爲分化低的,屬高度惡性。這種分級法雖有其優點,對臨牀治療和判斷預後也有一定意義。但缺乏定量標準,也不能排除主觀因素的影響。因此,如何建立精確的分級標準還待進一步研究。
腫瘤的分期目前有不同的方案,其主要原則是根據原發腫瘤的大小、浸潤的深度、範圍以及是否累及鄰近器官、有無局部和遠處淋巴結的轉移、有無血源性或其他遠處轉移等來確定腫瘤發展的程期或早晚。

惡性腫瘤細胞侵出基底膜進入細胞外間質,侵入血管並形成新的轉移竈(採自Basic Pathology)
腫瘤的分級和分期對臨牀上制定治療方案和估計預後有參考價值,特別是腫瘤的分期更爲重要,但是必須結合各種惡性腫瘤的生物學特性以及病人的全身情況等綜合考慮。
10 腫瘤對機體的影響
腫瘤因其良惡性的不同,對機體的影響也有所不同。
10.1 良性腫瘤
因其分化較成熟,生長緩慢,停留於局部,不浸潤,不轉移,故一般對機體的影響相對較小,主要表現爲局部壓迫和阻塞症狀。其影響的發生主要與其發生部位和繼發變化有關。如體表良性瘤除少數可發生的局部症狀外,一般對機體無重要影響;但若發生在腔道或重要器官,也可引起較爲嚴重的後果,如消化道良性腫瘤(如突入腸腔的平滑肌瘤),有時引起腸梗阻或腸套疊;顱內的良性瘤(如腦膜瘤、星形膠質細胞瘤)可壓迫腦組織阻塞腦室系統而引起顱內壓升高和相應的神經系統症狀。良性腫瘤有時可發生繼發性改變,亦可對機體帶來程度不同的影響,如腸的腺瘤性息肉、膀胱的乳頭狀瘤等表面可發生潰瘍而引起出血和感染。此外,內分泌腺的良性腫瘤則常因能引起某種激素分泌過多而產生全身性影響,如垂體前葉的嗜酸性腺瘤(acidophil adenoma)可引起巨人症(gigantism)或肢端肥大症(acromegaly);胰島細胞瘤分泌過多的胰島素,可引起陣發性血糖過低等。
10.2 惡性腫瘤
惡性腫瘤由於分泌不成熟、生長較快,浸潤破壞器官的結構和功能,並可發生轉移,因而對機體的影響嚴重。惡性瘤除可引起與上述良性瘤相似的局部壓迫和阻塞症狀外,發生於消化道者更易併發潰瘍、出血、甚致穿孔,導致腹膜炎,後果更爲嚴重。有時腫瘤產物或合併感染可引起發熱。腫瘤浸潤、壓迫局部神經還可引起頑固性疼痛等症狀。惡性腫瘤的晚期患者,往往發生惡病質(cachexia),可致患者死亡。惡病質是指機體嚴重消瘦、無力、貧血和全身衰竭的狀態,其發生機制尚未完全闡明,可能由於缺乏食慾,進食減少、出血、感染、發熱或因腫瘤組織壞死所產生的毒性產物等引起機體的代謝紊亂所致。此外,惡性腫瘤的迅速生長,消耗機體大量的營養物質,以及由於晚期癌瘤引起的疼痛,影響患者的進食及睡眠等,也是招致惡病質的重要因素。近年來發現巨噬細胞產生的腫瘤壞死因子(TNF)可降低食慾和增加分解代謝,與惡病質的發生有一定關係。
近年來相繼發現一些非內分泌腺腫瘤能產生和分泌激素或激素類物質,如促腎上腺皮質激素(ACTH)、甲狀旁腺素(PTH)、胰島素、抗利尿激素(ADH)、人絨毛膜促性腺激素(HCG)、促甲狀腺激素(TSH)、生長激素(GH)、降鈣素(calcitonin)等十餘種,可引起內分泌紊亂的臨牀症狀。此類腫瘤稱爲異位內分泌腫瘤,其所引起的臨牀症狀稱爲異位內分泌綜合徵。此類腫瘤大多爲惡性腫瘤,其中以癌爲多,如肺癌、胃癌、肝癌、胰腺癌、結腸癌等;也可見於肉瘤,如纖維肉瘤,平滑肌肉瘤、橫紋肌肉瘤和未分化肉瘤等。許多分泌異位激素的惡性腫瘤都有產生兩種以上激素的特點。關於異位激素產生的機制於今尚無一致的解釋,可能與瘤細胞內基因異常表達有關。除上述異位內分泌綜合徵外,由於腫瘤的產物或異常免疫反應(包括交叉免疫、自身免疫和免疫複合物沉着等)或其他不明原因,還可引起神經、消化、造血、骨關節、腎及皮膚等系統發生一些病變和臨牀表現。這些表現不是由原發腫瘤或轉移竈所在部位直接引起的,而是通過上述途徑間接引起,故稱爲副腫瘤綜合徵(paraneoplastic syndrome)。有時這些綜合徵可以表現得非常明顯而造成嚴重後果。認識此種綜合徵的意義在於它可能是一些隱匿腫瘤的早期表現,可由此而發現早期腫瘤。再者不要誤認爲這些系統的改變是由腫瘤轉移所致,而放棄對腫瘤的治療。與之相反,如腫瘤治療有效,這些綜合徵可減輕或消失。因此有十分重要的臨牀意義。
11 良性腫瘤與惡性腫瘤的區別
綜上所述,良性腫瘤和惡性腫瘤在生物學特點上是明顯不同的,因而對機體的影響也不同。良性腫瘤一般對機體影響小,易於治療,療效好;惡性腫瘤危害較大,治療措施複雜,療效還不夠理想。如果把惡性腫瘤誤診爲良性腫瘤,就會延誤治療或治療不徹底,造成復發、轉移。相反,如把良性腫瘤誤診爲惡性腫瘤,也必然要進行一些不必要、不恰當的治療,使患者遭受不應有的痛苦、損害和精神負擔。因此,區別良性腫瘤與惡性腫瘤,對於正確的診斷和治療具有重要的實際意義。現將良性腫瘤與惡性腫瘤的區別列於表7-1。
| 良性腫瘤 | 惡性腫瘤 | |
| 組織分化程度 | 分化好,異型性小,與原有組織的形態相似 | 分化不好,異型性大,與原有組織的形態差別大 |
| 核分裂 | 無或稀少,不見病理核分裂像 | 多見,並可見病理核分裂像 |
| 生長速度 | 緩慢 | 較快 |
| 生長方式 | 膨脹性和外生性生長,前者常有包膜形成,與周圍組織一般分界清楚,故通常可推動 | 浸潤性和外生性生長,前者無包膜,一般與周圍組織分界不清楚,通常不能推動,後者每伴有浸潤性生長 |
| 繼發改變 | 很少性發生壞死、出血 | 常發生出血、壞死、潰瘍形成等 |
| 轉移 | 不轉移 | 常有轉移 |
| 復發 | 手術後很少復發 | 手術等治療後較多復發 |
| 對機體影響 | 較小,主要爲局部壓迫或阻塞作用。如發生在重要器官也可引起嚴重後果 | 較大,除壓迫、阻塞外,還可以破壞原發處和轉移處的組織,引起壞死出血合併感染,甚至造成惡病質 |
必須指出,良性腫瘤與惡性腫瘤間有時並無絕對界限,有些腫瘤其表現可以介乎兩者之間,稱爲交界性腫瘤(如卵巢交界性漿液性乳頭狀囊腺瘤和粘液性囊腺瘤)。此類腫瘤有惡變傾向,在一定的條件下可逐漸向惡性發展。在惡性腫瘤中,其惡性程度亦各不相同,有的較早發生轉移,如鼻咽癌;有的轉移晚,如子宮體腺癌;有的則很少發生轉移。此外,腫瘤的良惡性也並非一成不變,有些良性腫瘤如不及時治療,有時可轉變爲惡性腫瘤,稱爲惡性變(malignant change),如結腸腺瘤性息肉可惡變爲腺癌。而個別的惡性腫瘤如黑色素瘤,有時由於機體免疫力加強等原因,可以停止生長甚至完全自然消退。又如見於少年兒童的神經母細胞瘤(neuroblastoma)的瘤細胞有時能發育成爲成熟的神經細胞,有時甚至轉移竈的瘤細胞也能繼續分化成熟,使腫瘤停止生長而自愈。但這種情況畢竟罕見,絕大多數惡性腫瘤能否逆轉爲良性,是目前腫瘤研究的重要課題之一。
12 腫瘤的命名與分類
12.1 腫瘤的命名原則
人體任何部位、任何組織、任何器官幾乎都可發生腫瘤,因此腫瘤的種類繁多,命名也複雜。一般根據其組織發生即組織來源來命名。良性腫瘤在其來源組織名稱後加“瘤”字,例如來源於纖維結締組織的良性瘤稱爲纖維瘤(fibroma),來源於腺上皮的良性瘤稱爲腺瘤(adenoma)等。有時還結合腫瘤的形態特點命名,如腺瘤呈乳頭狀生長並有囊腔形成者稱爲乳狀頭狀囊腺瘤。惡性腫瘤一般亦根據其組織來源來命名。來源於上皮組織的統稱爲癌(carcinoma),命名時在其來源組織名稱之後加“癌”字,如來源於鱗狀上皮的惡性腫瘤稱爲鱗狀細胞癌,來源於腺上皮呈腺樣結構的惡性腫瘤稱爲腺癌等。從間葉組織(包括纖維結締組織、脂肪、肌肉、脈管、骨、軟骨組織等)發生的惡性腫瘤統稱爲肉瘤(sarcoma),其命名方式是在來源組織名稱之後加“肉瘤”,例如纖維肉瘤、橫紋肌肉瘤、骨肉瘤等。惡性腫瘤的外形具有一定的特點時,則又結合形態特點而命名,如形成乳頭狀及囊狀結構的腺癌,則稱爲乳頭狀囊腺癌。如一個腫瘤中既有癌的結構又有肉瘤的結構,則稱爲癌肉瘤(carcinosarcoma)。
在病理學上,癌是指上皮組織來源的惡性腫瘤,但一般人所說的“癌症”(cancer),習慣上常泛指所有惡性腫瘤。
有少數惡性腫瘤不按上述原則命名,如有些來源於幼稚組織及神經組織的惡性腫瘤稱爲母細胞瘤,如神經母細胞瘤、髓母細胞瘤、腎母細胞瘤等;有些惡性腫瘤成分複雜或由於習慣沿襲,則在腫瘤的名稱前加“惡性”,如惡性畸胞瘤、惡性淋巴瘤、惡性黑色素瘤等。有些惡性腫瘤冠以人名、如尤文(Ewing)瘤、何傑金(Hodgkin)病;或按腫瘤細胞的形態命名,如骨的巨細胞瘤、肺的燕表細胞癌。至於白血病、精原細胞癌則是少數採用習慣名稱的惡性腫瘤,雖稱爲“瘤”或“病”,實際上都是惡性腫瘤。
12.2 腫瘤的分類
腫瘤的分類通常是以其組織發生(即來源於何種組織)爲依據,每一類別又分爲良性與惡性兩大類。茲舉例如下(表2)。
| 組織來源 | 良性腫瘤 | 惡性腫瘤 | 好發部位 |
| 一、上皮組織 | |||
| 鱗狀上皮 | 乳頭狀瘤 | 鱗狀細胞癌 | 乳頭狀瘤見於皮膚、鼻、鼻竇、喉等處;鱗癌見於宮頸、皮膚、食管、鼻咽、肺、喉和陰莖等處 |
| 基底細胞 | 基底細胞癌 | 頭面部皮膚 | |
| 腺上皮 | 腺瘤 | 腺瘤(各種類型) | 腺瘤多見於皮膚、甲狀腺、胃、腸;腺癌見於胃、腸、乳腺、甲狀腺等 |
| 粘液性或漿液性囊腺瘤 | 粘液性或漿液性囊腺癌 | 卵巢 | |
| 多形性腺瘤 | 惡性多形性腺瘤 | 涎腺 | |
| 移行上皮 | 乳頭狀瘤 | 移行上皮癌 | 膀胱、腎盂 |
| 二、間葉組織 | |||
| 纖維結締組織 | 纖維瘤 | 纖維肉瘤 | 四肢 |
| 纖維組織細胞 | 纖維組織細胞瘤 | 惡性纖維組織細胞瘤 | 四肢 |
| 脂肪組織 | 脂肪瘤 | 脂肪肉瘤 | 前者多見於皮下組織,後者多見於下肢和腹膜後 |
| 平滑組織 | 平滑肌瘤 | 平滑肌肉瘤 | 子宮和胃腸 |
| 橫紋肌組織 | 橫紋肌瘤 | 橫紋肌肉瘤 | 肉瘤多見於頭頸、生殖泌尿道及四肢 |
| 血管和淋巴管組織 | 血管瘤、淋巴管瘤 | 血管肉瘤 淋巴管肉瘤 | 皮膚和皮下組織、舌、脣等 |
| 骨組織 | 骨瘤 | 骨肉瘤 | 骨瘤多見於顱骨、長骨;骨肉瘤多見於長骨兩端,以膝關節上下尤爲多見 |
| 巨細胞瘤 | 惡性巨細胞瘤 | 股骨上下端、脛骨上端、肱骨上端 | |
| 軟骨組織 | 軟骨瘤 | 軟骨肉瘤 | 軟骨瘤多見於手足短骨;軟骨肉瘤多見於盆骨、肋骨、股骨、肱骨及肩胛骨等 |
| 滑膜組織 | 滑膜瘤 | 滑膜肉瘤 | 膝、踝、肩和肘等關節附近 |
| 間皮 | 間皮瘤 | 惡性間皮瘤 | 胸膜、腹膜 |
| 三、淋巴造血組織 | |||
| 淋巴組織 | 惡性淋巴瘤 | 頸部、縱隔、腸繫膜和腹膜後淋巴結 | |
| 造血組織 | 各種白血病 | 淋巴造血組織 | |
| 多發性骨髓瘤 | 椎骨、胸骨、肋骨、顱骨和長骨 | ||
| 四、神經組織 | |||
| 神經衣組織 | 神經纖維瘤 | 神經纖維肉瘤 | 單發性;全身皮神經;多發性:深部神經及內臟也受累 |
| 神經鞘細胞 | 神經鞘瘤 | 惡性神經鞘瘤 | 頭、頸、四肢等處神經 |
| 膠質細胞 | 膠質細胞瘤 | 惡性膠質細胞瘤 | 大腦 |
| 原始神經細胞 | 髓母細胞瘤 | 小腦 | |
| 腦膜組織 | 腦膜瘤 | 惡性腦膜瘤 | 腦膜 |
| 交感神經節 | 節細胞神經瘤 | 神經母細胞瘤 | 前者多見於縱隔和腹膜後,後者多見於腎上腺髓質 |
| 五、基他腫瘤 | |||
| 黑色素細胞 | 黑痣 | 惡性黑色素瘤 | 皮膚、粘膜 |
| 胎盤組織 | 葡萄胎 | 絨毛膜上皮癌、惡性葡萄胎 | 子宮 |
| 性索 | 支持細胞、間質細胞瘤 | 惡性支持細胞、間質細胞瘤 | 卵巢、睾丸 |
| 顆粒細胞瘤 | 惡性顆粒細胞瘤 | 卵巢 | |
| 生殖細胞 | 精原細胞瘤 | 睾丸 | |
| 無性細胞瘤 | 卵巢 | ||
| 胚胎性癌 | 睾丸、卵巢 | ||
| 三個胚葉組織 | 畸胎瘤 | 惡性畸胎瘤 | 卵巢、睾丸、縱隔和骶尾部 |
13 腫瘤的病因學和發病學
腫瘤病因學研究引起腫瘤的始動因素,腫瘤發病學則研究腫瘤的發病機制與腫瘤發生的條件。要治癒腫瘤和預防腫瘤的發生,關鍵問題是查明腫瘤的病因及其發病機制。
關於腫瘤的病因學和發病學,多年來進行了廣泛的研究,雖然至今尚未完全闡明,但近年來分子生物學的迅速發展,特別是對癌基因和腫瘤抑制基因的研究,已經初步揭示了某些腫瘤的病因與發病機制,例如伯基特淋巴瘤和人類T細胞白血病/淋巴瘤。目前的研究表明,腫瘤從本質上說是基因病。引起遺傳物質DNA損害(突變)的各種環境的與遺傳的致癌因子可能以協同的或者序貫的方式,激活癌基因或(和)滅活腫瘤的抑制基因,使細胞發生轉化(transformation)。被轉化的細胞可先呈多克隆性增生,經過一個漫長的多階段的演進過程(progression),其中一個克隆可相對無限制地擴增,通過附加突變,選擇性地形成具有不同特點的亞克隆(異質性),從而獲得浸潤和轉移的能力(惡性轉化),形成惡性腫瘤。圖7-19示腫瘤的病因和發病機制模式。

13.1 腫瘤發生的分子生物學基礎
1.癌基因
(1)原癌基因、癌基因及其產物:現代分子生物學的重大成就之一是發現了原癌基因(proto-oncogene)和原癌基因具有轉化成致癌的癌基因(oncogene)的能力。Bishop和Varmus因爲在這方面的貢獻而獲得1989年的諾貝爾獎。
癌基因是首先在逆轉錄病毒(RNA病毒)中發現的。含有病毒癌基因的逆轉錄病毒能在動物迅速誘發腫瘤並能在體外轉化細胞。後來在正常細胞的DNA中也發現了與病毒癌基因幾乎完全相同的DNA序列。被稱爲細胞癌基因。如ras,myc等。由於細胞癌基因在正常細胞中乃以非激活的形式存在,故又稱爲原癌基因。原癌基因可以由於多種因素的作用使其結構發生改變,而被激活成爲癌基因。
原癌基因編碼的蛋白質大多是對正常細胞生長十分重要的細胞生長因子和生長因子受體,如血小板衍生生長因子(PDGF)、纖維母細胞生長因子(FGF)、表皮細胞生長因子受體(EGF-R)、重要的信號轉導蛋白質(如酪氨酸激酶、絲氨酶-蘇氨酸激酶等)以及核調節蛋白(如轉錄激活蛋白)等。表4 示常見的癌基因及其產物。
| 編碼的蛋白質 | 原癌基因 | 激活機制 | 相關人類腫瘤 |
| 生長因子: | |||
| PDGF-β鏈 | sis | 過度表達 | 星形細胞瘤,骨肉瘤 |
| FGF | hst-1,int-2 | 過度表達 | 胃癌,膀胱癌,乳腺癌 |
| 生長因子受體: | |||
| EGF受體 | erb-B1 | 擴增 | 膠質瘤 |
| EGF樣受體 | neu(erb-B2) | 擴增 | 乳腺癌,卵巢癌,腎癌 |
| 信號轉導蛋白: | |||
| GTP-結合蛋白 | ras | 點突變 | 多種人體腫瘤,包括肺,結腸,胰,白血病 |
| 酪氨酸激酶 | abl | 易位 | 慢性粒細胞性,急性淋巴細胞性白血病 |
| 核調節蛋白: | |||
| 轉錄激活蛋白 | myc | 易位 | 伯基特淋巴瘤 |
| N-myc | 擴增 | 神經母細胞瘤,小細胞肺癌 | |
| 線粒體蛋白: | bcl-2 | 易位 | 濾泡性B-細胞淋巴瘤 |
(2)原癌基因的激活:原癌基因在各種環境的或遺傳的因素作用下,可發生結構改變(突變)而變爲癌基因;也可以是原癌基因本身結構沒有改變,而是由於調節原癌基因表達的基因發生改變使原癌基因過度表達。以上基因水平的改變可繼而導致細胞生長刺激信號的過度或持續出現,使細胞發生轉化。引起原癌基因突變的DNA結構改變包括點突變(如90%的胰腺癌有ras 基因的點突變)、染色體易位(如伯基特淋巴瘤的t(8:14),慢性粒細胞白血病的Phl染色體)、插入誘變、基因缺失和基因擴增(如神經母細胞瘤的N-myc原癌基因可複製成多達幾百個拷貝,在細胞遺傳學上表現爲染色體出現雙微小體和均染區)。癌基因編碼的蛋白質(癌蛋白)與原癌基因的正常產物相似,但有質或量的不同。通過生長因子或生長因子受體增加、產生突變的信號轉導蛋白與DNA結合的轉錄因子等機制,癌蛋白調節其靶細胞的代謝、促使該細胞逐步轉化,成爲腫瘤細胞。
2.腫瘤抑制基因 與原癌基因編碼的蛋白質促進細胞生長相反,在正常情況下存在於細胞內的另一類基因——腫瘤抑制基因的產物能抑制細胞的生長。若其功能喪失則可能促進細胞的腫瘤性轉化。由此看來,腫瘤的發生可能是癌基因的激活與腫瘤抑制基因的失活共同作用的結果。目前瞭解最多的兩種腫瘤抑制基因是Rb基因和P53基因。它們的產物都是以轉錄調節因子的方式控制細胞生長的核蛋白。其它腫瘤抑制基因還有神經纖維瘤病-1基因、結腸腺瘤性息肉基因、結腸癌丟失基因和Wilms瘤-1等。
(1)Rb基因:Rb基因隨着對一種少見的兒童腫瘤——視網膜母細胞瘤的研究而最早發現的一種腫瘤抑制基因。Rb基因的純合子性的丟失見於所有的視網膜母細胞瘤及部分骨肉瘤、乳腺癌和小細胞肺癌等。Rb基因定位於染色體13q14,編碼一種核結合蛋白質(P105-Rb)。它在細胞核中以活化的脫磷酸化和失活的磷酸化的形式存在。活化的Rb蛋白對於細胞從G0/G1期進入S期有抑制作用。當細胞受到刺激開始分裂時,Rb 蛋白被磷酸化失活,使細胞進入S期。當細胞分裂成兩個子細胞時,失活的(磷酸化的)Rb蛋白通過脫磷酸化再生使子細胞處於G1期或G0的靜止狀態。如果由於點突變或13q14的丟失而使Rb基因失活,則Rb蛋白的表達就會出現異常,細胞就可能持續地處於增殖期,並可能由此惡變。
(2)p53基因:p53基因定位於17號染色體。正常的p53蛋白(野生型)存在於核內,在脫磷酸化時活化,有阻礙細胞進入細胞週期的作用。在部分結腸癌、肺癌、乳腺癌和胰腺癌等均發現有p53基因的點突變或丟失,從而引起異常的p53蛋白表達,而喪失其生長抑制功能,從而導致細胞增生和惡變。近來還發現某些DNA病毒,例如HPV和SV-40,其致癌作用是通過它們的癌蛋白與活化的Rb蛋白或p53蛋白結合並中和其生長抑制功能而實現的。
3.多步癌變的分子基礎 惡性腫瘤的發生是一個長期的、多因素造成的分階段的過程,這已由流行病學、遺傳學和化學致癌的動物模式所證明。近年來的分子遺傳學研究從癌基因和腫瘤抑制基因的角度爲此提供了更加有力的證明。單個基因的改變不能造成細胞的完全惡性轉化,而是需要多基因的改變,包括幾個癌基因的激活和兩個或更多腫瘤抑制基因的喪失。以結腸癌的發生爲例,在從結腸上皮過度增生到結腸癌的演進過程中,關鍵性的步驟是癌基因以及腫瘤抑制基因的喪失或突變。這些階段梯性積累起來的不同基因分子水平的改變,可以在形態學的改變上反映出來(圖10)。

圖10 結直腸癌通過上皮增生-腺瘤-癌的階梯性的演進的分子生物學和形態學改變的關係(採自Kumar)
13.2 環境致癌因素及其致癌機制
化學致癌因素
現已確知的對動物有致癌作用的化學致癌物約有1000多種,其中有些可能和人類癌瘤有關。對化致癌物的研究表明:
①各種化學致癌物在結構上是多種多樣的。基中少數不需在體內進行代謝轉化即可致癌,稱爲直接作用的化學致癌物,如烷化劑。絕大多數則只有在體內(主要是在肝)進行代謝,活化後才能致癌,稱爲間接作用的化學致癌物或前致癌物,其代謝活化產物稱終末致癌物。如3,4-苯並芘是間接致癌物,其終末致癌物是環氧化物;
②所有的化學致癌物在化學上都具有親電子結構的基團,如環氧化物、硫酸酯基團等。它們都與細胞大分子的親核基團(如DNA分子中的鳥嘌呤的N-7、C-8,腺嘌呤的N-1、N-3,胞嘧啶的N-3等)共價結合,形成加合物,導致DNA的突變。化學致癌物大多數是致突變劑(mutagens);
③某些化學致癌物的致癌作用可由其它無致癌作用的物質協同作用而增大。這種增加致癌效應的物質稱爲促癌物(promoter),如巴豆油、激素、酚和某些藥物。致癌物引發初始變化稱爲激發作用(initiation),而促癌物的協同作用稱爲促進作用(promotion)。據此,Berenblum(1942)提出致癌過程的第二階段學說,即激發和促進兩個過程。現在認爲激發過程是由致癌物引起的不可逆的過程,使得一種原癌基因(如ras基因)突變性活化,這種突變可遺傳給子代細胞;促進過程據目前研究,可能是由於促癌劑(如巴豆油)是細胞內信號轉導通道的關鍵性成分——蛋白激酶C的活化劑,並且能使某些細胞分泌生長因子所致。因此促進作用能促使突變的細胞克隆性生長、抑制其正常分化,最後在附加突變的影響下形成惡性腫瘤。此學說在預惡性腫瘤具有現實意義,因爲激發過程是很短暫的,大多不可逆轉,而促進過程則很長,一般需10~20年。因此,如能減少環境中的促癌因子,亦可有效地預防惡性腫瘤的發生。
主要的化學致癌物質有以下幾類:
1)多環芳烴:存在於石油、煤焦油中。致癌性特別強的有3,4-苯並芘,1,2,5,6-雙苯並蒽,3-甲基膽蒽及9,10-二甲苯蒽等。這些致癌物質在使用小劑量時即能在實驗動物引起惡性腫瘤,如塗抹皮膚可引起皮膚癌,皮下注射可引起纖維肉瘤等。3,4-苯並芘是煤焦油的主要致癌成分,還可由於有機物的燃燒而產生。它存在於工廠排出的煤煙、菸草點燃後的煙霧中。近幾十年來肺癌的發生率之日益增加,公認與吸菸和工業城市嚴重的大氣污染有密切關係。此外,據調查,煙燻和燒烤的魚、肉等食品中也含有多環芳烴,這可能和某些地區胃癌的發病率較高有一定關係。多環芳烴在肝經細胞色素氧化酶P450系統氧化成環氧化物,後者以其親電子基因(不飽和的C-C鍵)與核酸分子以共價健結合而引起突變。
2) 芳香胺類與氨基偶氮染料:致癌的芳香胺類,如乙荼胺、聯苯胺、4-氨基聯苯等,與印染廠工人和橡膠工人的膀胱癌發生率較高有關。氨基偶氮染料,如以前在食品工業中曾使用過的奶油黃(二甲基氨基偶氮苯,可將人工奶油染成黃色的染料)和猩紅,在動物實驗可引起大白鼠的肝細胞性肝癌。以上兩類化學致癌物主要在肝代謝。芳香胺的活化是在肝通過細胞色素氧化酶P450系統使其N端羥化形成羥胺衍生物,然後與葡萄糖醛酸結合成葡萄糖苷酸從泌尿道排出,並在膀胱水解釋放出活化的羥胺而致膀胱癌。
3)亞硝胺類:亞硝胺類物質致癌譜很廣,可在許多實驗動物誘發各種不同器官的腫瘤。但是近年來引起很大興趣的主要是可能引起人體胃腸癌或其它腫瘤。亞硝酸鹽可作爲肉、魚類食品的保存劑與着色劑進入人體;也可由細菌分解硝酸鹽產生。在胃內的酸性環境下,亞硝酸鹽與來自食物的各種二級胺合成亞硝胺。我國河南林縣的流行病學調查表明,該地食管癌發病率很高與食物中的亞硝胺高含量有關。亞硝胺在體內經過羥化作用而活化,形成有很強的反應性的烷化碳離子而致癌。
4)真菌毒素:黃曲黴菌廣泛存在於高溫潮溼地區的黴變食品中,尤以黴變的花生、玉米及穀類含量最多。黃麴黴毒素有許多種,其中黃麴黴毒素B1(aflatoxin B1)的致癌性最強,據估計其致癌強度比奶油黃大900倍,比二甲基亞硝胺大75倍,而且化學性很穩定,不易被加熱分解,煮熟後食入仍有活性。黃麴黴毒素B1的化學結構爲異環芳烴,在肝通過肝細胞內的混合功能氧化酶氧化成環氧化物而致突變。這種毒素主要誘發肝細胞性肝癌。我國和南非肝癌高發區的調查都顯示黃麴黴毒素B1在食物中的污染水平與肝癌的發病率有關。但這些地區同時也是乙型肝炎病毒(HBV)感染的高發區。在HBV感染與黃麴黴毒素B1污染之間的關係方面,分子生物學的研究表明,黃麴黴毒素B1的致突變作用是使腫瘤抑制基因p53發生點突變而失去活性,而HBV感染所致的肝細胞慢性損傷和由此引起的肝細胞持續再生爲黃麴黴毒素B1的致突變作用提供了有利的條件。因此HBV感染與黃麴黴毒素B1的協同作用是我國肝癌高發地區的主要致癌因素。此外,也已證明,在我國食管癌高發地區居民食用的酸菜中分離出的白地黴菌,其培養物有促癌或致癌作用。
(2)直接作用的化學致癌物:這類化學致癌物不需要體內代謝活化即可致癌,一般爲弱致癌劑,致癌時間長。
1)烷化劑與酰化劑:例如抗癌藥中的環磷酰胺、氮芥、本丁酸氮芥、亞硝基脲等。這類具有致癌性的藥物可在應用相當長時間以後誘發第二種腫瘤,如在化學治療痊癒或已控制的白血病、何傑金淋巴瘤和卵巢癌的病人,數年後可能發生第二種腫瘤,通常是粒細胞性白血病。某些使用烷化劑的非腫瘤病人,如類風溼性關節炎和Wegener肉芽腫的病人,他們發生惡性腫瘤的機率大大高於正常人。因此這類藥物應謹慎使用。
2)其它直接致癌物:金屬元素對人類也有致癌的作用,如鎳、鉻、鎘、鈹等,如煉鎳工人中,鼻癌和肺癌明顯高發;鎘與前列腺癌、腎癌的發生有關;鉻可引起肺癌等。其原因可能是金屬的二價陽離子,如鎳、鎘、鉛、鈹、鈷等,是親電子的,因此可與細胞大分子,尤其是DNA反應。例如鎳的二價離子可以使多聚核苷酸解聚。一些非金屬元素和有機化合物也有致癌性,如砷可誘發皮膚癌;氯乙烯可致塑料工人的肝血管肉瘤;苯致白血病等,也受到關注。
化學致癌大多與環境污染和職業因素有關,因此徹底治理環境污染,防治職業病,對於減少惡性腫瘤的發病是極其重要的。目前發現的具有防癌或抗癌作用的稀有元素有鉬、硒、鎂、鉑等。表5列出主要的化學致癌物、易感人羣和誘發的腫瘤。
| 化學致癌物 | 易感人羣 | 誘發的主要腫瘤 |
| 直接作用的 | ||
| 烷化劑 | 接受化學治療的惡性腫瘤病人 | 白血病 |
| 間接作用的 | ||
| 多環芳烴 | 吸菸者、食用熏製魚肉者 | 肺癌、胃癌 |
| 芳香胺 | 染料工人、橡膠工人 | 膀胱癌 |
| 亞硝胺 | 亞硝酸鹽污染食物的食用者 | 食管癌、胃癌 |
| 黃麴黴毒素B1 | 污染食物的食用者 | 腫細胞性腫癌 |
| 石棉纖維 | 礦工、接觸者 | 肺癌、胸膜間皮瘤 |
| 氯乙烯 | 塑料廠工人 | 肝血管肉瘤 |
| 苯 | 橡膠工人、染料工人 | 白血病 |
| 砷 | 礦工、農藥工人和噴撒者 | 皮膚癌、肺癌、肝癌 |
| 鎳 | 煉鎳工人 | 鼻癌、肺癌 |
| 鉻 | 接觸含鉻氣體者 | 鼻癌、肺癌、喉癌 |
| 鎘 | 接觸者 | 前列腺癌、腎癌 |
物理性致癌因素
已證實的物理性致癌因素主要是離子輻射。異物、慢性炎性刺激和創傷亦可能與促癌有關。
(1)電離輻射:包括X射線、γ射線、亞原子微粒(β粒子、質子、中子或α粒子)的輻射以及紫外線照射。大量事實證明,長期接觸X射線及鐳、鈾、氡、鈷、鍶等放射性同位素,可以引起各種不同的惡性腫瘤,例如放射工作者長期接觸X射線而又無必要的防護措施時,常可發生手部放射性皮炎以致皮膚癌;其急性和慢性粒細胞性白血病的發生率亦較一般人高10倍以上。在出生前或出生後接受過X線照射的兒童,其急性白血病的發生率高於一般兒童。開採含放射性物質(鈷、氡等)的礦工易患肺癌。日本長崎、廣島在第二次世界大戰時受原子彈爆炸影響的倖存居民,經過長期觀察,發現慢性粒細胞白血病的發生率明顯增高(照射後4~8年爲發病高峯),甲狀腺癌、乳腺癌、肺癌等的發生率亦較高。在嬰幼兒期接受過頸部放射線照射者,甲狀腺癌發生率明顯增高。有些放射性同位素如32P、48Sr、210Po、239Pu等攝入能誘發骨肉瘤。
輻射能使染色體斷裂、易位和發生點突變,因而激活癌基因或者滅活腫瘤抑制基因。由於與輻射有關的腫瘤的潛伏期較長,因此腫瘤最終可能當輻射所損傷的細胞的後代又受到其它環境因素(如化學致癌劑、病毒等)所致的附加突變之後,纔會出現。
動物實驗和臨牀觀察均證實,陽光中紫外線長期過度照射可引起外露皮膚的鱗狀細胞癌、基底細胞癌和惡性黑色素瘤。白種人或照射後色素不增加的有色人種最易發生。其作用機制是細胞內DNA吸收了光子,使其中相鄰的兩個嘧啶連接(包括胸腺嘧啶與胸腺嘧啶、胸腺嘧啶與胞嘧啶、胞嘧啶與胞嘧啶),形成嘧啶二聚體。二聚體又形成環丁烷,從而破壞DNA雙螺旋中二聚體所在處的磷酸二酯骨架,妨礙DNA分子的複製。在正常人這種損害通常可爲一系列DNA修復酶所修復,因此皮膚癌發病少見。而一種罕見的常染色體隱性遺傳病——着色性幹皮病(xeroderma pigmentosum)的患者,由於先天性缺乏修復DNA所需的酶,不能將紫外線所致的DNA的損害修復,皮膚癌的發病率很高。
(2)熱輻射的促癌作用:克什米爾人冬季習慣用懷爐取暖,有時在腹部引起“懷爐癌”;我國西北地區居民冬季燒火取暖,有時臀部皮膚發生癌變形成所謂“炕癌”。這些說明長期的熱輻射可能有一定的促癌作用。在燒傷瘢痕的基礎上易發生“瘢痕癌”,有人在燒傷瘢痕中發現化學致癌物。
(3)慢性炎性刺激:腫瘤必須在細胞增生的基礎上發生。慢性炎症時產生的細胞生長因子能使細胞持續增生,在此基礎上DNA易發生突變而發生腫瘤。因而慢性刺激有促癌作用。慢性皮膚潰瘍、結石引起的慢性膽囊炎、慢性子宮頸炎和子宮內膜增生等病變有時可發生癌變,可能即與此有關。
(4)異物:石棉和石棉製品能導致人的胸膜間皮瘤,重度暴露於石棉纖維的工人,其胸膜間皮瘤的發生率可達2%~3%,潛伏期一般爲20年。動物實驗證明,植入動物體內的異物,如塑料、金屬片,玻璃纖維等,可以誘發各種肉瘤。腫瘤的發生與植入物體的化學性關係不大而與物體表面的形狀、光滑程度和耐久性有關。人類對於異物刺激有較大的耐受性,雖然尚無植入塑料和金屬致癌的報道,但是已人工乳房(有機硅橡膠製成)與乳腺癌有關的報告。
另一種與腫瘤有關的異物是寄生蟲。醫學文獻中早就有埃及血吸蟲感染的病人,其膀胱癌的發生率高於正常人的報告。
RNA病毒(逆轉錄病毒)的研究導致了癌基因的發現,並由此開創了腫瘤的分子遺傳學。在人類已知的與腫瘤有關的病毒並不多。
(1)RNA致瘤病毒:對動物逆轉錄病毒致癌的研究發現,由於病毒類型的不同,它們是通過轉導(transduction)或插入突變(insertional mutagenesis)這兩種機制將其遺傳物質整合到宿主細胞DNA中,並使宿主細胞分生轉化的。①急性轉化病毒:這類病毒含有從細胞的原癌基因轉導的病毒癌基因,如src,abl,myb等,這些病毒感染細胞後,將以其病毒RNA爲模板通過逆轉錄酶合成的DNA片斷整合(integration)到宿主的DNA鏈中並表達,導致細胞的轉化;②慢性轉化病毒:這類病毒(如鼠乳腺癌病毒)本身並不含有癌基因,但是有促進基因,當感染宿主細胞後促進基因也可由於逆轉錄酶的作用而插入到宿主細胞DNA鏈中的原癌基因附近,引起正常的或突變的原癌基因激活並且過度表達,使宿主細胞轉化。
人類T細胞白血病/淋巴瘤病毒I(human T-cell leukemia/lymhoma virus I,HTVL-1)是與人類腫瘤發生密切相關的一種RNA病毒,與主要流行於日本和加勒比地區的T細胞白血病/淋巴瘤有關。HTLV-1病毒與AIDS病毒一樣,轉化的靶細胞是CD4+的T細胞亞羣(輔助T細胞)。HTLV-1在人類是通過性交、血液製品和哺乳傳播的。受染人羣發生白血病的機率爲1%,潛伏期爲20~30年。HTLV-1轉化T細胞的機制還不甚清楚。HTLV-1不含有任何已知的癌基因,也未發現其在某一原癌基因附近的固定的整合位置。HTLV-1的轉化活性與其RNA中的一個稱爲Tax的基因有關。Tax的產物對病毒的複製十分重要,因其通過對5’-長末端重複片段(5’-long terminal repeat region,5’-LTR)的作用刺激病毒mRNA的轉錄。Tax蛋白也可激活幾種能引起T細胞增生的宿主基因的轉錄,如編碼調節細胞內其它基因表達的P55蛋白c-fos基因,編碼PDGF的c-sis基因,編碼IL-2及其受體的基因和髓樣生長因子(即粒-單核細胞集落刺激因子,GM-CSF)的基因。IL-2及其受體的基因激活後可以建立起一個自分泌體系(autocrine system)能直接引起T細胞的增生;GM-CSF作用於巨噬細胞,使其產生IL-1,從而引起T細胞的增生。因此HTLV-1是通過Tax基因轉化細胞的。這些增生的T細胞最初是多克隆性的,而且出現二次突變的可能性大大增加,如其中的某一個發生第二次突變,將導致單克隆性的T細胞腫瘤。
(2)DNA致瘤病毒:DNA病毒中有50多種可引起動物腫瘤。對它們的研究,尤其是對多瘤病毒的研究,提示了DNA病毒致癌的機制。DNA病毒感染細胞後出現兩種後果:①如果病毒DNA未能被整合到宿主的基因組中,病毒的複製不會受到干擾,大量的病毒複製最終使細胞死亡;②要引起細胞的轉化,病毒基因必需整合到宿主的DNA中並且作爲細胞的基因加以表達。多瘤病毒的T基因編碼的蛋白質T抗原具有酪氨酸激酶活性,能像生長因子受體那樣刺激細胞DNA合成,並使細胞持續增生,而後形成腫瘤。與人類腫瘤發生密切有關的DNA病毒有以下三種:
1)人類乳頭狀瘤病毒(human papilloma virus,HPV):HPV與人類上皮性腫瘤,主要是與子宮頸和肛門生殖器區域的鱗狀細胞癌的關係,近年來已有大量資料予以證實。HPV的某些亞型(如16、18型)的DNA序列已在75%~100%的宮頸癌病例的癌細胞中發現。HPV的致癌機制還不完全清楚。近來發現HPV的16、18和31高危亞型的早期病毒基因產物E6和E7蛋白,極易與Rb和p53基因的產物結合並中和其抑制細胞生長的功能。在體外,Rb和p53基因產物的失活能使人類棘細胞轉化並且長期存活,但不形成腫瘤。這時如果再轉染一個突變的ras基因,就會引起完全的惡性轉化。這說明HPV在致癌時不是單獨作用的,需要環境因素的協同。
2)Epstein-Barr病毒(EBV):與之有關的人類腫瘤是伯基特淋巴瘤和鼻咽癌。
伯基特淋巴瘤是一種B細胞性的腫瘤。流行於非洲東部和散發於世界各地。在流行地區,所有病人的瘤細胞都攜帶EBV的基因組成分並且出現特異的染色體易位t(8:14)。EBV對B細胞有很強的親和性,能使受染的B細胞發生多克隆性的增生。在正常的個體這種增生是可以控制的,受染者沒有症狀或者臨牀表現爲自限性的傳染性單核細胞增生症。而在非洲流行區,由於瘧疾或其它感染損害了患者的免疫功能,受染B細胞乃持續增生。在此基礎上如再發生附加的突變(如t(8:14)),則後者使c-myc激活,導致進一步的生長控制喪失,並在其它附加基因損傷的影響下,最終導致單克隆性的腫瘤出現。
鼻咽癌在我國南方和東南亞流行,EBV的基因組也在幾所有腫瘤細胞中發現。但是與伯基特淋巴瘤一樣,EBV在鼻咽癌發生中的作用也需要其它因素的配合。
3)乙型肝炎病毒(hepatitis b virus,HBV):流行病學調查發現,慢性HBV感染與肝細胞性肝癌的發生有密切的關係。但是HBV本身並不含有可以編碼任何轉化蛋白(癌蛋白)的基因,其中肝細胞DNA中的整合也沒有固定的模式。HBV的致癌作用看來是多因素的:
①如在前文所述,HBV導致的慢性肝損傷使肝細胞不斷再生,這使另外的致癌因素(如黃麴黴毒素B1)的致突變作用容易發生;
14 影響腫瘤發生、發展的內在因素及其作用機制
腫瘤發生和發展是一個十分複雜的問題,除了外界致癌因素的作用外,機體的內在因素也起着重要作用,後者包括宿主對腫瘤反應,以及腫瘤對宿主的影響。這些內在因素是複雜的,許多問題至今尚未明瞭,還有待進一步研究。機體的內在因素可分爲以下幾方面;
遺傳因素
人類腫瘤是否有遺傳性,以及遺傳因素在人類腫瘤的發生上究竟起多大的作用?這是人們普遍關注並不斷深入研究的課題。根據一些高癌家族系譜的分析,有以下幾種不同情況:
(1)呈常染色顯性遺傳的腫瘤:如視網膜母細胞瘤、腎母細胞瘤、腎上腺或神經節的神經母細胞瘤等。一些癌前疾病,如結腸多發性腺瘤性息肉症、神經纖維瘤病等本身不是惡性腫瘤,但惡變率極高,有100%的結腸家族性多發性腺瘤性息肉病的病例在50歲以前發生惡變,成爲多發性結腸腺癌。這些腫瘤和癌前疾病都屬單基因遺傳,以常染色體顯性遺傳的規律出現。其特點爲早年(兒童期)發病,腫瘤呈多發性,常累及雙側器官。
(2)呈常染色體隱性遺傳的遺傳綜合徵:如患Bloom綜合徵(先天性毛細血管擴張性紅斑及生長發育障礙)時易發生白血病及其他惡性腫瘤;毛細血管擴張性共濟失調症患者多發生急性白血病和淋巴瘤;着色性幹皮病患者經紫外光照射後易患皮膚基底細胞癌、磷狀細胞癌或黑色素瘤。這些腫瘤易感性高的人常伴有某種遺傳缺陷,如免疫缺陷、染色體缺陷和內切酶等的缺陷。
(3)遺傳因素與環境因素在腫瘤發生中起協同作用,而環境因素更爲重要,決定這類腫瘤的遺傳因素是屬於多基因的。目前發現不少常見腫瘤有家族史,如乳腺癌、胃腸癌、食管癌、肝癌、鼻咽癌、白血病、子宮內膜癌、前列腺癌、黑色素瘤等。
總的說來不同的腫瘤可能有不同遺傳傳遞方式,真正直接遺傳的只是少數不常見的腫瘤。遺傳因素在大多數腫瘤發生中的作用是對致癌因子的易感性或傾向性。Kundson(1974)提出二次突變假說(two hit hypothesis)來解釋遺傳性損害在腫瘤發生中作用。以現代分子生物學的術語來描述這一假說是:以視網膜母細胞瘤例,Rb基因定位於染色體13q14,只有兩條同源染色體上的Rb等位基因都被滅活,即需經兩次突變後,才能使腫瘤發生。在家族性視網膜母細胞瘤患兒基因組中已經存在一個從父母得到的有缺陷的Rb基因拷貝,另一個Rb基因拷貝正常的(雜合型),因而只要再有一次體細胞突變,即可形成腫瘤(純合型)。這種家族性視網膜母細胞瘤的患兒年齡小,雙側發病的較多。而在散發性的視網膜母細胞瘤的患兒,由於其兩個正常的Rb等位基因都要通過體細胞突變而失活才能發病,故出現這種病例的可能性只有家族性的萬分之一,而且發病較晚,多爲單側。
腫瘤免疫惡性轉化是由於遺傳基因的改變引起的。有些異常基因表達的蛋白可引起免疫系統的反應,從而使機體能消滅這些“非已”的轉化細胞。如果沒有這種免疫監視機制,則腫瘤的發生要比實際上出現的多得多。關於腫瘤免疫的研究不僅對腫瘤的發生有重要的意義。而且爲腫瘤的免疫治療指出了方向。
(1)腫瘤抗原:引起機體免疫反應的腫瘤抗原可分爲兩類:①只存在於腫瘤細胞而不存在於正常細胞的腫瘤特異性抗原;②存在於腫瘤細胞和某些正常細胞的腫瘤相關抗原。
儘管在腫瘤特異性抗原的研究上花費了大量的時間和精力,企圖尋找某種腫瘤的特異性抗原。但是現已在化學致癌的動物模型中發現,腫瘤特異性抗原是個體獨特的,即不同個體中同一種致癌物誘發的同一組織學類型的腫瘤有不同的特異性抗原。因此用檢測某種腫瘤特異性抗原來診斷或用某抗體來治療某些腫瘤的可能性在目前尚不存在。腫瘤特異性抗原的個體獨特性的原因是,癌變時癌基因發生突變的隨機性引起異常蛋白的隨機出現,因而無法產生特定的針對某一類腫瘤的抗原。
腫瘤相關抗原在腫瘤中的表達,推測與遺傳因素的改變有關。它們又可分爲兩類:腫瘤胚胎抗原和腫瘤分化抗原。前者在正常情況下出現在發育中的胚胎組織而不見於成熟組織,但可見於癌變組織。例如在胚胎肝細胞和肝細胞性肝癌中出現的甲胎蛋白以及在胚胎組織和結腸癌中出現的癌胚抗原。後者是指腫瘤細胞具有的與分化程度有關的某些抗原。例如前列腺特異抗原見於正常前列腺上皮和前列腺癌細胞。腫瘤相關抗原在有關腫瘤的診斷上是有用的標記,也可用此製備抗體,用於腫瘤的免疫治療。
(2)抗腫瘤的免疫效應機制:腫瘤免疫反應以細胞免疫爲主,體液免疫爲輔。參加細胞免疫的效應細胞主要有細胞毒性T細胞(CTL)、自然殺傷細胞(NK)和巨噬細胞。抗腫瘤免疫的機制如圖11所示。CTL被白細胞介素2(IL-2)激活後可以通過其T細胞受識別瘤細胞上的人類主要組織相容性複合體(major histocompatibility compolex,MHC)I型分子而釋放某些溶解酶將瘤細胞殺滅。CTL的保護作用在對抗病毒所致的腫瘤(如EBV引起的伯基特淋巴瘤和HPV導致的腫瘤)時特別明顯。NK細胞是不需要預先致敏的、能殺傷腫瘤細胞的淋巴細胞。由IL-2激活後,NK細胞可以溶解多種人體腫瘤細胞,其中有些並不引起T細胞的免疫反應,因此NK細胞是抗腫瘤免疫的第一線的抵抗力量。NK細胞識別靶細胞的機制可能是通過NK細胞受體和抗體介導的細胞毒作用(antibody-dependent cellular cytotoxicity,ADCC)。巨噬細胞在抗腫瘤反應中是與T細胞協同作用的:T細胞產生的α-干擾素可激活巨噬細胞,而巨噬細胞產生的腫瘤壞死因子(TNF-α)和活性氧化代謝產物在溶解瘤細胞中起主要作用。此外巨噬細胞的Fc受體還可與腫瘤細胞表面的IgG結合,通過ADCC殺傷腫瘤細胞。體液免疫參加抗腫瘤反應的機制主要是激活補體和介導NK細胞參加的ADCC。

CTL:細胞毒性T淋巴細胞 NK:自然殺傷細胞 IFN-γ:γ-干擾素 TNF-α:腫瘤壞死因子α IL-2:白細胞介素2 MHCI:主要組織相容性複合體Ⅰ型分子
(3)免疫監視:免疫監視機制在抗腫瘤中的作用的最有力的證據是,在免疫缺陷病患者和接受免疫抑制治療的病人中,惡性腫瘤的發病率明顯增加。先天性免疫缺陷病(如X-性聯無γ球蛋白血癥)的病人有5%發生惡性腫瘤,這比對照組高出200倍。在器官移植的受者和AIDS患者中發生淋巴瘤的可能也大大增加。惡性腫瘤患者隨着病程的發展和病情惡化每伴有免疫功能普遍下降,這在晚期患者尤爲突出。相反,有些腫瘤,如神經母細胞瘤、惡性黑色素瘤和絨毛膜上皮癌等腫瘤患者,由於機體免疫功能增高,腫瘤可發生自發消退。但大多數惡性腫瘤乃發生於免疫功能正常的人羣,這些腫瘤能逃脫免疫系統的監視並破壞機體的免疫系統,其機制還不甚清楚,可能與下列因素有關:①在腫瘤生長過程中,具有較強抗原性的亞克隆被免疫系統消滅,而無抗原性的或者抗原性弱的亞克隆則生長成腫瘤;②CTL攻擊腫瘤細胞時要識別瘤細胞膜上的Ⅰ型MHC抗原,故腫瘤細胞MHC抗原表達的喪失或減少,會使瘤細胞避開CTL的攻擊;③許多致癌因素同時也抑制宿主的免疫反應,如化學致癌物和電離輻射等;④腫瘤產物也可以抑制免疫反應,如由許多腫瘤分泌的轉化生長因子β(TGF-β)就是一種潛在免疫抑制劑;⑤腫瘤引發的有些免疫反應,如抑制性T細胞的激活,本身就可抑制對腫瘤的免疫反應。
(4)腫瘤的免疫治療:免疫治療旨在通過替換免疫系統中的受到抑制的成分,或者刺激內源性的反應來增加機體的抗腫瘤能力。目前有三種主要的方法。①繼承性細胞治療:先將病人的周圍血淋巴細胞與IL-2一起孵育,然後將產生的淋巴激肽激活殺傷細胞(lymphokine-activated killer cell,LAK細胞,主要由NK細胞組成)再輸回病人;或者將從病人切除的腫瘤組織中的淋巴細胞(主要是CTL)在體外與IL-2一起培養後再輸回病人。這兩種治療方法的效果還有待於評價;②細胞激肽治療:主要是用IL-1,IFN-α、γ、TNF-α等細胞激肽與其它治療方法一起用於抗癌,目前療效最好的是INF-α對毛細胞白血病的治療;③抗體治療:目前的興趣集中在用抗腫瘤相關抗原的單克隆抗體與細胞毒素結合後製成的“免疫毒素”來治療腫瘤,這種“魔彈”的療效正在觀察中。
其它與腫瘤發病學有關的因素
(1)內分泌因素:內分泌紊亂與某些器官腫瘤的發生發展有密切的關係,如乳腺癌的發生發展可能與患者體內雌激素水平過高或雌激素受體的異常有關。乳腺癌在妊娠期和哺乳期發展得特別快,切除卵巢或用雌激素治療可使腫瘤明顯縮小。此外,激素與惡性腫瘤的轉移及擴散也有一定的關係,如垂體前葉激素可促進腫瘤的生長和轉移;腎上腺皮質激素對某些造血系統的惡性腫瘤有抑制其生長和轉移的作用。
(2)性別和年齡因素:腫瘤的發生在性別上有很大的差異,除生殖器官腫瘤及乳腺癌在女性明顯較多見外,膽囊、甲狀腺及膀胱等器官的腫瘤也是女性多於男性。肺癌、食管癌、肝癌、胃癌、鼻咽癌和結腸癌等則以男性爲多見。性別上的這種差異,其原因除一部分與女性激素有關外,主要是可能與男女的性染色體的不同和某一性別較多地接受某種致癌因子的作用有關。
年齡在腫瘤的發病上也有一定的意義。一般來說,腫瘤的發生隨年齡的增大而增加,這可用體細胞突變累積來解釋。而多見於幼兒和兒童的腫瘤,常與遺傳性的基因損害有關,如視網膜母細胞瘤、腎母細胞瘤和神經母細胞瘤等。
(3)種族和地理因素:某些腫瘤在不同種族或地區中的發生率有相當大的差別,如歐美國家的乳腺癌年死亡率是日本的4~5倍,而日本的胃癌年死亡率比美國高7倍。在我國廣東、四川和香港、新加坡等地廣東人中,鼻咽癌相當常見而且發病年齡較輕。這說明腫瘤與種族有一定的關係。但是也有移民材料說明移居美國的華僑和日僑中,胃癌的發生率在第三代已有明顯的下降。因此地理和生活習慣可能也起到一定的作用。
綜上所述,隨着分子生物學的發展,近年來對於腫瘤的病因與發病機制的研究有了很大的進展。但是腫瘤的發生發展是異常複雜的,目前瞭解的只是一角,還有許多未知的領域。但以下幾點是迄今比較肯定的:
③環境的和遺傳的致癌因素引起的細胞遺傳物質(DNA)改變的主要靶基因是原癌基因和和腫瘤抑制基因。原癌基因的激活和(或)腫瘤抑制基因的失活可導致細胞的惡性轉化。